Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.

Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
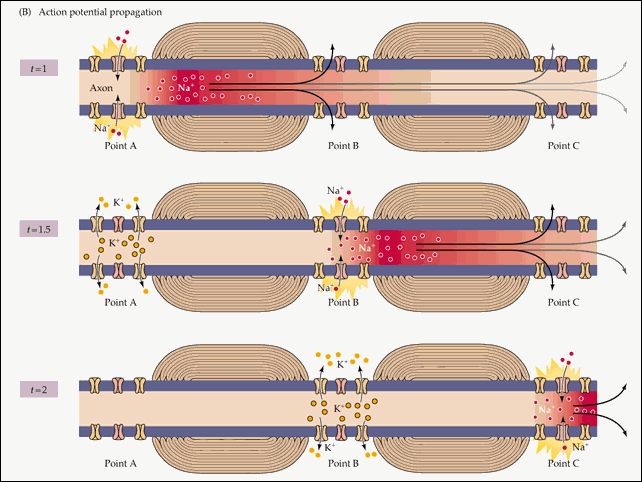
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.

3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.
8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.
Нервный импульс - это движущаяся волна изменений в состоянии мембраны. Она включает в себя структурные изменения (открытие и закрытие мембранных ионных каналов), химические (изменяющиеся трансмембранные потоки ионов) и электрические (изменения электрического потенциала мембраны: деполяризацию, позитивную поляризацию и реполяризацию). © 2012-2019 Сазонов В.Ф..
Можно сказать короче:
"Нервный импульс - это волна изменений, движущаяся по мембране нейрона". © 2012-2019 Сазонов В.Ф..
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Потенциал действия - это электрическая характеристика (электрическая составляющая) нервного импульса.
Нервный импульс - это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия - это только электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно). (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Электрические явления – развитие потенциала действия.
Химические явления – движение ионных потоков.
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением . Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация , т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
«овершут» – положительная поляризация (инверсия, или реверсия, заряда мембраны);
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Образно можно сказать, что заряженные ионы – это и есть создатели электрических потенциалов в нервных клетках. Для многих людей звучит странно утверждение, что вода не проводит электрический ток. Но на самом деле это так. Сама по себе вода является диэлектриком, а не проводником. В воде электрический ток обеспечивают не электроны, как в металлических проводах, а заряженные ионы: положительные катионы и отрицательные анионы. В живых клетках основную «электрическую работу» выполняют катионы, так как они более подвижны. Электрические токи в клетках – это потоки ионов.
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками . Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы .
На молекулярно-структурном уровне происходит открытие мембранных натриевых каналов. Сначала этот процесс идет пропорционально силе внешнего воздействия, а затем становится просто «неудержимым» и массовым. Открытие каналов обеспечивает вход натрия в клетку и вызывает деполяризацию. Затем, примерно через 2-5 миллисекунд, происходит их автоматическое закрытие . Это закрытие каналов резко обрывает движение ионов натрия внутрь клетки, и, следовательно, обрывает нарастание электрического потенциала. Рост потенциала прекращается, и на графике мы видим спайк. Это вершина кривой на графике, дальше процесс пойдет уже в обратном направлении. Конечно, очень интересно разобраться в том, что натриевые каналы имеют двое ворот, и открываются они активационными воротами, а закрываются инактивационными, но это следует обсуждать ранее, в теме «Возбуждение». Мы на этом останавливаться не будем.
Параллельно в открытием натриевых каналов с небольшим отставанием во времени идет нарастающее открытие калиевых каналов. Они медлительные по сравнению с натриевыми. Открытие дополнительных калиевых каналов усиливает выход положительных ионов калия из клетки. Выход калия противодействует «натриевой» деполяризации и вызывает восстановление полярности (восстановление электроотрицательности). Но натриевые каналы опережают калиевые, они срабатывают примерно в 10 раз быстрее. Поэтому входящий поток положительных ионов натрия в клетку опережает компенсирующий выход ионов калия. И поэтому деполяризация развивается опережающими темпами по сравнению с противодействующей ей поляризацией, вызванной утечкой ионов калия. Вот почему, пока натриевые каналы не закроются, восстановление поляризации не начнется.
Пожар как метафора распространяющегося возбуждения
Для того чтобы перейти к пониманию смысла динамичного процесса возбуждения, т.е. к пониманию его распространения вдоль мембраны, надо представить себе, что описанные нами выше процессы захватывают сначала ближайшие, а затем все новые, все более и более отдаленные участки мембраны, пока не пробегут по всей мембране полностью. Если вы видели «живую волну», которую устраивают болельщики на стадионе за счет вставания и приседания, то вам легко будет представить себе мембранную волну возбуждения, которая образуется за счет последовательного протекания в соседних участках трансмембранных ионных токов.
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
Можно ли «погасить» волну распространяющегося возбуждения?
Вряд ли нервная клетка способна «погасить» этот начавшийся «пожар» возбуждения. Поэтому первый способ подходит только для искусственного вмешательства в работу нейронов (например, в лечебных целях). Но вот «залить водичкой» некоторые участки и поставить блок распространению возбуждения, оказывается, вполне возможно.
© Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии - деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью - невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Потенциал действия - волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляетэлектрический разряд - быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.
Потенциал действия развивается на мембране в результате её возбуждения и сопровождается резким изменением мембранного потенциала.
В потенциале действия выделяют несколько фаз:
Фаза деполяризации;
Фаза быстрой реполяризации;
Фаза медленной реполяризации (отрицательный следовый потенциал);
Фаза гиперполяризации (положительный следовый потенциал).
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциал чувствительных Na+- каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации. В результате деполяризации мембраны происходит открытие потенциалчувствительных К+- каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется. Усиливает реполяризацию поступление в клетку Ca2+ Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+ помпы. Поступление в клетку Cl– дополнительно гиперполяризует мембрану Изменение величины мембранного потенциала во время развития потенциала действия связано в первую очередь с изменением проницаемости мембраны для ионов натрия и калия.
Современные представления о механизме его генерации
Методом фиксации мембранного потенциала удалось измерить токи, текущие через плазмолемму аксона (аксолемму) кальмара и убедиться в том, что в покое ток катионов (К +) направлен из цитоплазмы в интерстиций, а при возбуждении доминирует ток катионов (Na +) в клетку. В состоянии «покоя» плазмолемма почти непроницаема для ионов, находящихся в межклеточном пространстве(Na + С1 - и НСОз - ,).
При возбуждении проницаемость для ионов натрия на время, равное нескольким миллисекундам, резко возрастает, а затем снова падает. В результате катионы (ионы Na +) и анионы (С1 - , НСОз) разобщаются на плазмолемме: Na + входит в цитоплазму, а анионы нет. Поток положительных зарядов в цитоплазму не только компенсирует потенциал покоя, но и превышает его. Возникает так называемый «овершут» (или инверсия мембранного потенциала). Входящий поток натрия - результат его пассивного движения по открывшимся мембранным каналам по концентрационному и электрическому градиентам. Выходящий поток этого катиона обеспечивается калий-натриевой помпой.
Виды электрических ответов (электротонический потенциал, локальный ответ, потенциал действия). Механизм их возникновения.
В пpоцессе pазвития возбуждения плазматической мембpаны (изменения ее ионной пpоницаемости и электpического состояния) в зависимости от силы pаздpажителя возникает тpи вида электpических ответов:
Электpотонический потенциал
Локальный ответ
Потенциал действия
Электpотонический потенциал
Электротонический потенциал - это пассивный сдвиг величины мембранного потенциала (МП) при действии подпорогового стимула электрического тока.
1. Возникает в ответ на действие катода постоянного тока по силе воздействия меньше 0,5 поpоговой величины
2. Сопpовождается пассивной, слабо выpаженной электpотонической деполяpизацией за счет "-" заpяда катода (ионная пpоницаемость мембpаны пpактически не изменяется), котоpая наблюдается только во вpемя действия pаздpажителя
3. Развитие и исчезновение потенциала пpоисходит по экспоненциальной кpивой и опpеделяется паpаметpами
4. pаздpажающего тока, а также сопpотивлением и емкостью мембpаны
5. Такой вид возбуждения имеет местный хаpактеp и не может pапpспpостpаняться
6. Увеличивает возбудимость ткани
Механизм возникновения
Простейшая модель раздражимости при прохождении тока представляет собой процесс, при котором положительные заряды тока кратковременно разряжают, т.е. деполяризуют мембрану, что вызывает нарушение равновесия ионных потоков.
Во время деполяризации больше ионов калия (+К) покидает клетку и тем самым уравновешивается поток ионного и электрического тока, что, в свою очередь, приводит к стабилизации заряда мембранной емкости. Сдвиг потенциала, вызываемый импульсом тока, называетсяэлектротоническим потенциалом, илиэлектротоном .
Скорость нарастания электротонического потенциала определяется в основном емкостью мембраны. Однако большинство нервных клеток имеют вытянутую форму. Нервное волокно иногда достигает длины 1 м при диаметре 1 мкм. Следовательно, выходя из такой клетки, пропускаемый через нее ток будет распределяться очень неравномерно. Установлено, что по мере увеличения расстояния от источника возбуждения (тока) временной ход электротонического потенциала (электротона) постепенно замедляется. Происходит это потому, что электротон преодолевает сопротивление не только мембраны, но продольное сопротивление внутренней среды самой нервной клетки. Для малых сдвигов потенциала электротонические потенциалы в нерве можно зарегистрировать на расстоянии не более нескольких сантиметров от места их возникновения, т.е. локально.
Деполяризующий электротонический потенциал, который превышает пороговый уровень, вызывает возбуждение. Возбуждение возможно тогда, когда импульс тока имеет адекватную длительность и амплитуду. Соответственно определенный уровень длительности и амплитуды импульса тока существенно влияет на передачу информации в форме потенциала действия. В этой связи локальный характер деполяризации дендиритов, тел нервных клеток и аксонов различается.
Деполяризация дендритов и соответственно тел нервных клеток наблюдается едва достигается пороговый уровень. Происходит это потому, что деполяризация идет за счет повышения натриевой (+Nа) проницаемости мембраны, которая в дальнейшем продолжает деполяризацию автоматически.

Локальный ответ
Локальный потенциал (ЛП) - это местное нераспространяющееся подпороговое возбуждение, существующее в пределах от потенциала покоя (-70 мВ в среднем) до критического уровня деполяризации (-50 мВ в среднем). Его длительность может быть от нескольких миллисекунд до десятков минут.
1. Возникает в ответ на действие pаздpажителя силой от 0,5 до 0,9 поpога
2. Активная фоpма деполяpизации, поскольку ионная пpоницаемость повышается в зависимости от силы подпоpогового pаздpажителя
3. Гpадуален по амплитуде (амплитуда находится в пpямой зависимости от силы и частоты pаздpажений)
4. Развитие деполяpизации пpоисходит до кpитического уpовня, пpичем не пpямолинейно, а по S-обpазной кpивой. Пpи этом деполяpизация пpодолжает наpастать после пpекpащения pаздpажения, а затем сpавнительно медленно исчезает
5. Способен к суммации (пpостpанственной и вpеменной)
6. Локализуется в пункте действия pаздpажителя и пpактически не способен к pаспpостpанению, т.к. хаpактеpизуется большой степенью затухания
7. Повышает возбудимость стpуктуpы
Виды Локальных ответов(потенциалов):
1. Рецепторный. Возникает на рецепторных клетках (сенсорных рецепторах) или рецепторных окончаниях нейронов под действием стимула (раздражителя). Механизм возникновения такого рецепторного локального потенциала детально рассмотрен на примере восприятия звука слуховыми рецепторами - Молекулярные механизмы рецепции (трансдукции) звука по пунктам Этот процесс называется "трансдукция", то есть преобразование раздражения в нервное возбуждение. Сенсорные рецепторы вторичного типа не умеют порождять нервный импульс, поэтому их возбуждение остаётся локальным и от его амплитуды зависит то, сколько рецепторная клетка выбросит медиатора.
2. Генераторный . Возникает на сенсорных афферентных нейронах (на их дендритных окончаниях, перехватах Ранвье и/или аксонных холмиках) под действием медиаторов, которые выделили сенсорные клеточные рецепторы вторичного типа. Генераторный потенциал превращается в потенциал действия и нервный импульс при достижении им критического уровня деполяризации, т.е. он генерирует (порождает) нервный импульс. Потому он и назван генераторным.
3. Возбуждающий постсинаптический потенциал (ВПСП) . Возникает на постсинаптической мембране синапса, т.е. он отражает передачу возбуждения от одного нейрона к другому. Обычно он составляет +4 мВ. Важно отметить, что возбуждение передаётся от одного нейрона другому именно в виде ВПСП, а не готового нервного импульса. ВПСП вызывает деполяризацию мембраны, но подпороговую, не достигающую КУД и не способную породить нервный импульс. Поэтому обычно требуется целая серия ВПСП для того, чтобы родился нервный импульс, т.к. величина единичного ВПСП совершенно недостаточна для того, чтобы достичь критического уровня деполяризации. Вы можете сами подсчитать, сколько требуется одновременно действующих ВПСП, чтобы родился нервный импульс. (Ответ: 5-6.)
4. Тормозный постсинаптический потенциал (ТПСП) . Возникает на постсинаптической мембране синапса, но только не возбуждает её, а, наоборот, тормозит. Соотвтетственно, эта постсинаптическая мембрана входит в состав тормозного синапса , а не возбуждающего. ТПСП вызывает гиперполяризацию мембраны, т.е. сдвигает потенциал покоя вниз, подальше от нуля. Обычно он составляет -0,2 мВ. Используются два механизма создания ТПСП: 1) "хлорный" - происходит открытие ионных каналов для хлора (Cl-), через них в клетку входят ионы хлора и увеличивают её электроотрицательность, 2)"калиевый" - происходит открытие ионных каналов для калия (К+), через них выходят ионы калия, уносят из клетки положительные заряды, что увеличивает электроотрицательность в клетке.
5. Пейсмекерные потенциалы - это эндогенные близкие к синусоидальным периодические колебания мембранного потенциала с частотой 0,1-10 Гц и амплитудой 5-10 мВ. Их генерируют у себя специальные нейроны-пейсмекеры (водителями ритма) самостоятельно, без внешнего воздействия. Пейсмекерные локальные потенциалы обеспечивают периодическое достижение нейроном-пейсмекером критического уровня деполяризации и спонтанную (т.е. самопроизвольную) генерацию им потенциалов действия и, соответственно, нервных импульсов.
Механизм возникновения
Важно понять то, что процесс рождения локального потенциала начинается с открытия ионных каналов . Открытие ионных каналов - это самое главное! Их нужно открыть для того, чтобы в клетку пошёл поток ионов и принёс в неё электрические заряды. Эти ионные электрические заряды как раз и вызывают смещение электрического потенциала мембраны вверх или вниз, т.е. локальный потенциал.
натрия (Na+) , то в клетку вместе с ионами натрия попадают положительные заряды, и её потенциал смещается вверх в сторону нуля. Это - деполяризация, и так рождается возбуждающий локальный потенциал . Можно сказать, что возбуждающие локальные потенциалы порождаются натриевыми ионными каналами, когда они открываются.
Образно можно сказать и так: "Каналы открываются - потенциал рождается".
Если открываются ионные каналы для хлора (Cl-) , то в клетку вместе с ионами хлора попадают отрицательные заряды, и её потенциал смещается вниз ниже потенциала покоя. Это гиперполяризация, и таким способом рождается тормозный локальный потенциал . Можно сказать, что тормозные локальные потенциалы порождаются хлорными ионными каналами .
Существует также ещё один механизм формирования тормозных локальных потенциалов - за счёт открытия дополнительных ионных каналов для калия (К+) . В этом случае из клетки через них начинают выходить "лишние" порции ионов калия, они выносят положительные заряды и увеличивают электроотрицательность клетки, т.е. вызывают её гиперполяризацию. Таким образом, можно сказать, что тормозные локальные потенциалы порождаются дополнительными калиевыми ионными каналами .
Как видите, всё очень просто, главное - открыть нужные ионные каналы . Стимул-управляемые ионные каналы открываются раздражителем (стимулом). Хемо-управляемые ионные каналы открываются медиатором (возбуждающим или тормозным). Точнее, в зависимости от того на какие каналы (натриевые, калиевые или хлорные) будет действовать медиатор, таков будет и локальный потенциал - возбуждающий или тормозный. А медиатор как для возбуждающих локальных потенциалов, так и для тормозных, может быть одним и тем же, тут важно, какие ионные каналы будут связываться с ним своими молекулярными рецепторами - натриевые, калиевые или хлорные.
Потенциал действия
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
1. Возникает пpи действие pаздpажителей поpоговой и свеpхпоpоговой силы (может возникать пpи суммации подпоpоговых pаздpажителей вследствии достижения уpовня кpитической деполяpизации)
2. Активная деполяpизация пpотекает пpактически мгновенно и pазвивается пофазно (деполяpизация, pеполяpизация)
3. Hе имеет гpадуальной зависимости от силы pаздpажителя и подчиняется закону "все или ничего". Амплитуда зависит только от свойств возбудимой ткани
4. Hе способен к суммации
5. Снижает возбудимость ткани
6. Распpостpаняется от места возникновения по всей мембpане возбудимой клетки без изменения амплитуды
Механизм возникновения
Фаза деполяризации . Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
6. Возбудимость. Изменение возбудимости в процессе возбуждения.
А. Возбудимость клетки во время ее возбуждения быстро и сильно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и, так же как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов. Схематично эти изменения представлены на рис. 3.6.б.
1. Кратковременное повышение возбудимости в начале развития ПД, когда уже возникла частичная деполяризация клеточной мембраны. Если деполяризация не достигает критической величины, то регистрируется локальный потенциал. В случае, если деполяризация достигает Екр, то развивается ПД. При замедленном развитии начальной деполяризации она оценивается как препотенциал. Возбудимость повышена потому, что клетка частично деполяризована, мембранный потенциал приближается к критическому уровню, поскольку открывается часть потенциалчувствительных быстрых Na-каналов. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла Е кр, при которой возникает ПД.
2. Абсолютная рефракторная фаза - это полная невозбудимость клетки (возбудимость равна нулю), она соответствует пику ПД и продолжается 1-2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефракторная фаза. Клетка в этот период при любой силе раздражения не отвечает. Невозбудимость клетки в фазу деполяризации и инверсии (в первую ее половину - восходящая часть пика ПД) объясняется тем, что потенциалзависимые т -ворота Na-каналов уже открыты и ионы Na + быстро поступают в клетку по всем каналам. Те ворота Na-каналов, которые еще не успели открыться, открываются под влиянием деполяризации - уменьшения мембранного потенциала. Поэтому дополнительное раздражение клетки относительно движения ионов Na + в клетку ничего изменить не может.


Рис. 3.6. Фазовые изменения возбудимости клетки(б) во время ПД (а). 1,4 - возбудимость повышена; 2 - абсолютная рефрактерная фаза;
2. Относительная рефрактерная фаза - это период восстановления возбудимости, когда сильное раздражение может вызвать новое возбуждение (см. рис. 3.6,5, кривая 3). Относительная рефрактерная фаза соответствует конечной части фазы реполяризации от уровня Е кр ± 10 мВ и следовой гиперполяризации клеточной мембраны, что является следствием все еще повышенной проницаемости для ионов К + и избыточного выхода ионов К + -каналов из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, так как часть Nа + -каналов в конце реполяризации находится еще в состоянии инактивации, а выход ионов К + из клетки препятствует ее деполяризации. Кроме того, в период следовой гиперполяризации мембранный потенциал больше и, естественно, дальше отстоит от критического уровня деполяризации. Если реполяризация в конце пика ПД замедляется (см. рис. 3.6,а), то относительная рефрактерная фаза включает и период замедления реполяризации, и период гиперполяризации. Рис. 3.6.Фазовые изменениявозбудимости клетки (b) во времяПД (а).1,4-возбудимость повышена;2-абсолютная рефрактерная фаза;3-относительная рефрактерная фаза
4. Фаза экзальтации - это период повышенной возбудимости. Он соответствует следовой деполяризации. В нейронах ЦНС вслед за гиперполяризацией возможна частичная деполяризация клеточной мембраны. В эту фазу очередной ПД можно вызвать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Nа + . Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность.
Б. Лабильность, или функциональная подвижность (Н.Е.Введенский)- это скорость протекания одного цикла возбуждения, т.е. ПД. Как видно из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая, в свою очередь, зависит от скорости изменения проницаемости клеточной мембраны. Особое значение при этом имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани.
Биологическая реакция
– ответная реакция клеток, тканей, органов в ответ на раздражитель (стимул).
Раздражимость
– свойство всех живых тканей изменять своё внутреннее состояние при изменении внешних условий.
Виды тканей в зависимости от реагирования на внешние раздражители:
I Возбудимые
- обладают свойством возбудимости, т.е. способностью возбуждаться нервная, мышечная, железистая.
II Невозбудимые
– изменяют свое состояние, но не генерируют процесс возбуждения в ответ на нанесенный стимул.
Возбудимость
– способность ткани переходить в возбуждённое состояние.
Возбуждение
– деятельное состояние тканей в ответ на действие раздражителя, это сложная биологическая реакция, проявляющаяся в совокупности физических, физико-химических и функциональных изменений, способная к распространению по ткани.
Возбуждение включает в себя неспецифические и специфические компоненты.
Неспецифические:
сдвиг химических реакций, образование тепла, физико-химические изменения,
продукция биопотенциалов, структурные изменения в мембране клеток.
Специфические:
мышечная ткань отвечает мышечным сокращением, нервная ткань - генерацией нервного импульса и его проведением, железистая ткань – образованием и выделением секрета.
Возбуждение может быть локальным и динамическим (распространяющимся).
Биопотенциалы
Луиджи Гальвани 1791 г. в эксперименте показал, что живые ткани содержат «животное электричество», его научный оппонент, физик Вольта - что это электричество от разнородных металлов, он создал первый источник постоянного тока, который носит название гальванический элемент.
Виды биопотенциалов:
1. Биопотенциал покоя (мембранный) - МПП.
2. Биопотенциал действия (возбуждения) - ПД.
- Биопотенциал покоя – это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. Наружная поверхность мембраны клетки имеет положительный заряд, а внутренняя – отрицательный.
Биопотенциал покоя регистрируется внутриклеточным методом – с помощью микроэлектродов, один из которых вводится внутрь клетки (рис.1).
Рисунок 1. Схематическое представление метода регистрации биопотенциалов.
В эксперименте биопотенциал покоя можно зарегистрировать между повреждённым и неповреждённым участком ткани. Повреждённый участок является моделью внутренней поверхности мембраны клетки.
При внутриклеточном отведении перезарядка мембраны регистрируется под одним электродом (однофазный ПД), при внеклеточном отведении потенциал действия проходит через два электрода (регистрируется двухфазный ПД).
- Биопотенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения (рис.2). Измеряется ПД с помощью внутриклеточного отведения и внеклеточного отведения.

Рисунок 2. Потенциал действия, основные его фазы.
Современная, экспериментально доказанная, мембранно-ионная теория возникновения биопотенциалов (Ходжкин, Хаксли, Катц).
Основные положения:
- Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы (рис.3).
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала.

Рисунок 3. Строение биологической мембраны.
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала (рис.4).

Рисунок 4. Строение ионного канала.
Каналы для натрия имеют два типа ворот: быстрые активационные и медленные инактивационные. В покое открыты медленные инактивационные и закрыты быстрые активационные. При возбуждении происходит открытие быстрых активационных и медленное закрытие медленных инактивационных, т.е. на короткий промежуток времени оба типа ворот открыты (рис.5).

Рисунок 5. Работа активационные и инактивационный воротных механизмов натриевого ионного канала.
Калиевые каналы
имеют только медленные ворота.
Насосы выполняют функцию транспорта через мембрану ионов против градиента концентрации, для их работы используется энергия АТФ.
- По обе стороны мембраны существует концентрационный градиент.
Внутри клетки в 40 раз > К+; t;/p>
Вне клетки: в 20-30 раз > Na+,
в 50 раз > Cl-.
- Мембрана пропускает молекулы жирорастворимых веществ, а анионы органических кислот не проходят. Мембрана проницаема для воды, для ионов проницаемость мембраны различна: для калия в состоянии покоя проницаемость почти в 25 раз больше, чем для натрия. При возбуждении увеличивается проницаемость и для калия (постепенно), и для натрия (быстро, но на очень короткий промежуток времени).
Потенциал покоя
Проницаемость мембраны для ионов К+ повышена, поэтому калий играет основную роль в генерации МПП. Калий создаёт электрическое поле и заряжает наружную поверхность мембраны «+». В тот момент, когда «+» потенциал наружной стороны достигает определённой величины по отношению к «–» внутри, который создается анионами – наступает динамическое равновесие между входящими и выходящими из клетки ионами К+. Этому моменту соответствует потенциал равновесия для К - потенциал покоя.
МПП характеризуется:
1. постоянством;
2. полярностью, снаружи «+», внутри «-»;
3. величина – в мВ, для скелетной мышцы - 60 – 90 мВ,
для гладкой - -30 – 70 мВ,
для нерва -50 – 80мВ,
для секреторной клетки - -20мВ.
МПП - один из основных показателей состояния физиологического покоя клетки. При увеличении внеклеточной концентрации калия уменьшается МПП, т.к. уменьшается диффузия калия из клетки в связи со снижением его концентрационного градиента. При действии веществ, блокирующих ресинтез АТФ, т.к. прекращается работа натрий-калиевого насоса, также снижается МПП. Ионы натрия и хлора входят в клетку, но ввиду низкой проницаемости значительного влияния на МП не оказывают.
Потенциал действия
При возбуждении – резко увеличивается (в несколько тысяч раз) проницаемость для ионов Na, которые поступают внутрь клетки лавинообразно и заряжают внутреннюю сторону «+» - происходит деполяризация мембраны, а затем количество ионов натрия внутри превышает калиевый заряд на поверхности и это приводит к перезарядке мембраны (реверсии). Постепенно увеличивающаяся проницаемость для калия и его поток из клетки инактивирует натриевую проницаемость и приводит к восстановлению заряда на мембране. Возникает фаза реполяризации.
Существенным фактором является натрий-калиевый насос, который выводит из клетки 3 иона натрия в обмен на 2 иона калия, вводимые в клетку. Его работа зависит от метаболизма клетки, в частности, от ее энергоснабжения. При этом расходуется 1 молекула АТФ (рис.6).

Рисунок 6. Механизм работы натрий-калиевого насоса.
ПД состоит из пикового потенциала, который образуется фазой деполяризации, реверсии и реполяризации, и следовых потенциалов (рис.2).
Следовые потенциалы:
Отрицательный (следовая деполяризация);
Положительный (следовая гиперполяризация).
Причиной следовых потенциалов являются дальнейшие изменения соотношения между входом натрия в клетку и выходом калия из нее. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. При следовой гиперполяризации – остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса.
ПД характеризуется:
1. изменяющимся характером;
2. кратковременностью – несколько мсек;
3. зарядом мембраны, снаружи – «-», внутри – «+».
При действии веществ, блокирующих натриевые каналы, ПД не генерируется, т.к. в норме деполяризация мембраны обусловлена повышение ее натриевой проницаемости. При увеличении силы раздражителя выше порога амплитуда ПД не изменяется, т.к. не изменяется число активированных натриевых каналов, которые максимально раскрываются при пороговом раздражении.
Условия, необходимые для возникновения возбуждения (законы раздражения).
Возбудимость тканей различна. Чтобы вызвать возбуждение, раздражитель должен обладать:
1. Достаточной силой – закон порога.
2. Крутизной (градиентом) нарастания этой силы – закон аккомодации.
3. Временем действия – закон силы-времени.
1. Закон силы.
Мерой возбудимости является порог раздражения – минимальная сила раздражителя, способная вызвать возбуждение. Все раздражители можно разделить на подпороговые, пороговые и сверхпороговые. По биологическому значению раздражители делят на адекватные (действующие на ткань в естественных условиях, к ним она приспособлена в процессе эволюции) и неадекватные. В физиологических экспериментах в качестве раздражителя чаще всего используется электрический ток, т.к. он вызывает обратимые изменения, легко дозируется по силе и длительности, по своей природе близок к электрическим процессам, протекающим в живых организмах.
В 1870 г. Боудич в эксперименте на мышце сердца путем нанесения на неё одиночных пороговых раздражений регистрировал ответную реакцию - установил, что на подпороговое раздражение реакции не было, при пороговой силе и сверхпороговой амплитуда ответной реакции была одинаковой. На основании этого он предложил закон «Всё или ничего».
После введения в экспериментальные исследования микроэлектронной техники было установлено, что на подпороговое раздражение в ткани возникает ответная реакция. Если сила стимула меньше 50% пороговой величины, то под полюсами электродов происходит пассивная деполяризация без изменения проницаемости мембраны для ионов (электротонические изменения). Если сила стимула меньше пороговой величины, но больше 50% от нее, то в ткани возникает локальный ответ, который сопровождается деполяризацией мембраны в области нанесения раздражения и не распространяется на всю ткань, возбудимость тканей в этом участке повышена. Локальный ответ подчиняется закону силовых отношений, т.е. чем больше сила подпорогового стимула, тем больше амплитуда локального ответа. Проницаемость мембраны клетки в этом участке повышается для ионов натрия. При нанесении порогового стимула возникает ПД, амплитуда которого не изменяется, если величина стимула будет превышать пороговую, т.е. отвечает закону «Все или ничего», но на сверхпороговые стимулы длительность ПД будет меньше за счет укорочения продолжительности локального ответа.
Момент перехода локального ответа в ПД называется критическим уровнем деполяризации (КУД), а сдвиг заряда мембраны с мембранного потенциала до КУД, называется пороговым потенциалом, он наряду с порогом раздражения характеризует возбудимость ткани.
Изменение возбудимости тканей при возбуждении.
При возбуждении возбудимость тканей претерпевает определенные изменения в зависимости от фаз ПД (рис.7):
I – супернормальная возбудимость (первичная) соответствует локальному ответу, при этом два подпороговых стимула, нанесенных с интервалом времени, короче длительности локального ответа могут суммироваться и вызывать ПД;
II – абсолютная рефрактерность – соответствует регенеративной деполяризации и реверсии, при этом ткань становится абсолютно невозбудимой и не отвечает на самые сильные раздражители;
III – относительная рефрактерная фаза, соответствует реполяризации, при этом возбудимость ткани постепенно восстанавливается и сверхпороговый стимул, нанесенный в этот период может генерировать ПД;
IV – супернормальная возбудимость (вторичная или экзальфационная фаза) - следовой деполяризации, ткань становится более возбудимой, чем в исходном состоянии и даже подпороговый стимул способен вызвать ПД;
V – субнормальная возбудимость – следовой гиперполяризации, возбудимость ткани несколько снижена.

Рисунок 7. Изменение возбудимости мембраны при развитии потенциала действия.
2. Закон градиента раздражения (Дюбуа Реймон).
Чем больше градиент раздражения, тем больше (до известных пределов) реакция живого образования.
За время действия медленно нарастающего стимула наступает приспособление ткани – аккомодация. Она связана с тем, что при возбуждении проницаемость для ионов натрия увеличивается на короткий промежуток времени, если в течение его раздражитель не достигает пороговой величины, то увеличивающаяся проницаемость для ионов калия инактивирует натриевую проницаемость и возбуждение не наступает. При этом происходит также сдвиг КУД с увеличением порогового потенциала.
3. Закон силы-времени (Лапик).
Пороговая величина любого раздражителя находится в обратной зависимости от времени его действия, которая характеризуется математической кривой – гиперболой. Характер кривой свидетельствует о том, что подпороговые стимулы (меньше 1 реобазы) не вызовут возбуждение как долго бы они не действовали, в то же время очень сильный кратковременный стимул, длительность которого меньше полезного времени, также не вызовет возбуждение.
Сила постоянного тока, которая, действуя неопределенное время, вызывает возбуждение, называется реобазой.
Время, в течение которого ток в 1 реобазу вызывает возбуждение – полезное время.
Минимальное время, в течение которого ток силой в 2 реобазы вызовет возбуждение, называется хронаксией. Исследование этого показателя используется в неврологической и травматологической практике для изучения динамики восстановления в нервной или мышечной ткани после травмы.
Список использованной литературы
- Нормальна фізіологія /Під ред. В.І. Філімонова. – К. – Здоров’я, 1994. – С. 5 - 37.
- Физиология человека /Под ред. Г. И. Косицкого. – М., Медицина, 1985. – С. 19 – 84.
- Посібник з нормальної фізіології /Під ред. В.Г. Шевчука. – К., Здоров’я, 1995. – С. 6 - 36.
- Руководство к практическим занятиям по физиологии /Под ред. Г. И. Косицкого. – М., Медицина, 1988. – С. 72 - 94.
- Нормальная физиология /Под ред. В. И. Филимонова. - Запорожье, 1995. – С. 74-72.
- Физиология человека. Т.1 /Под ред. Р. Шмидта и Г. Тевса. – М., Мир, - 1996. - С. 9-87.
- Физиология человека. Т.1 / Под ред. В.М. Покровского. – М., Медицина, 1998. – С. 27-97.
- Общий курс физиологии человека и животных. Т.1. /Под ред. А.Д. Ноздрачева – М., Высшая школа, 1991.- С.36-116.
- Физиология человека. /Под ред. В.М. Смирнова – М., Медицина, 2002. – С. 45-61, 82-94.
- Фізіологія людини. Вільям Ф. Ганонг. – БаК, Львів, 2002. – С. 6 – 69, 74-76.
- управляемые. По механизму управления: электро-, хемо- и механоуправляемые;
- неуправляемые. Не имеют воротного механизма и всегда открыты, ионы идут постоянно, но медленно.
Потенциал покоя — это разность электрических потенциалов между наружной и внутренней средой клетки.
Механизм формирования потенциалов покоя. Непосредственная причина потенциала покоя — это неодинаковая концентрация анионов и катионов внутри и вне клетки. Во-первых, такое расположение ионов обосновано разницей проницаемости. Во-вторых, ионов калия выходит из клетки значительно больше, чем натрия.
Потенциал действия — это возбуждение клетки, быстрое колебание мембранного потенциала вследствие диффузии ионов в клетку и из клетки.
При действии раздражителя на клетки возбудимой ткани сначала очень быстро активируются и инактивируются натриевые каналы, затем с некоторым опозданием активируются и инактивируются калиевые каналы.
Вследствие этого ионы быстро диффундируют в клетку или из нее согласно электрохимическому градиенту. Это и есть возбуждение. По изменению величин и знака заряда клетки выделяют три фазы:
- 1-я фаза — деполяризация. Уменьшение заряда клетки до нуля. Натрий движется к клетке согласно концентрационному и электрическому градиенту. Условие движения: открыты ворота натриевого канала;
- 2-я фаза — инверсия. Изменение знака заряда на противоположный. Инверсия предполагает две части: восходящую и нисходящую.
Восходящая часть. Натрий продолжает двигаться в клетку согласно концентрационному градиенту, но вопреки электрическому градиенту (он препятствует).
Нисходящая часть. Калий начинает выходить из клетки согласно концентрационному и электрическому градиенту. Открыты ворота калиевого канала;
- 3-я фаза — реполяризация. Калий продолжает выходить из клетки согласно концентрационному, но вопреки электрическому градиенту.
Критерии возбудимости
При развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а, следовательно, исходное состояние возбудимой клетки. Это нормальный уровень возбудимости. Период предспайка — период самого начала потенциала действия. Возбудимость ткани слегка повышена. Эта фаза возбудимости — первичная экзальтация (первичная супернормальная возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения этого уровня сила раздражителя может быть меньше пороговой.
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны. Абсолютная рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются.
После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня — это фаза относительной рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов.
Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя исходной поляризации, то порог раздражения снижен, т.е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал увеличивается — возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
Механизм возникновения мембранного потенциала покоя
Каждая клетка в состоянии покоя характеризуется наличием трансмембранной разности потенциалов (потенциала покоя). Обычно разность зарядов между внутренней и внешней поверхностями мембран составляет от -80 до -100 мВ и может быть измерена с помощью наружного и внутриклеточного микроэлектродов (рис. 1).
Разность потенциалов между наружной и внутренней сторонами мембраны клетки в состоянии ее покоя называют мембранным потенциалом (потенциалом покоя).
Создание потенциала покоя обеспечивается двумя основными процессами — неравномерным распределением неорганических ионов между внутри- и внеклеточным пространством и неодинаковой проницаемостью для них клеточной мембраны. Анализ химического состав вне- и внутриклеточной жидкости свидетельствует о крайне неравномерном распределении ионов (табл. 1).
В состоянии покоя внутри клетки много анионов органических кислот и ионов К+, концентрация которых в 30 раз больше, чем снаружи; ионов Na+, наоборот, снаружи клетки в 10 раз больше, чем внутри; СI- также больше снаружи.
В покое мембрана нервных клеток наиболее проницаема для К+, менее — для СI- и очень мало проницаема для Na+/ Проницаемость мембраны нервного волокна для Na+ B покое в 100 раз меньше, чем для K+. Для многих анионов органических кислот мембрана в покое совсем непроницаема.
Рис. 1. Измерение потенциала покоя мышечного волокна (А) с помощью внутриклеточного микроэлектрода: М — микрозлектрод; И — индифферентный электрод. Луч на экране осциллографа (В) показывает, что до прокола мембраны микроэлектродом разность потенциалов между М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена отрицательно по отношению к ее наружной поверхности (по Б.И. Ходорову)
Таблица. Внутри- и внеклеточные концентрации ионов мышечной клетки теплокровного животного, ммоль/л (по Дж. Дудел)
|
Внутриклеточная концентрация |
Внеклеточная концентрация |
|
|
А- (анионы органических соединений) |
В силу градиента концентраций К+ выходит на наружную поверхность клетки, вынося свой положительный заряд. Высокомолекулярные анионы не могут следовать за К+ из-за непроницаемости для них мембраны. Ион Na+ также не может возместить ушедшие ионы калия, ибо проницаемость мембраны для него значительно меньше. СI- по градиенту концентраций может перемешаться только внутрь клетки, увеличивая тем самым отрицательный заряд внутренней поверхности мембраны. Вследствие такого перемещения ионов возникает поляризация мембраны, когда наружная ее поверхность заряжается положительно, а внутренняя — отрицательно.
Электрическое поле, которое создастся на мембране, активно вмешивается в распределение ионов между внутренним и наружным содержимым клетки. По мере возрастания положительного заряда на наружной поверхности клетки иону К+ как положительно заряженному становится все труднее перемещаться изнутри наружу. Он движется как бы в гору. Чем больше величина положительного заряда на наружной поверхности, тем меньшее количество ионов К+ может выходить на поверхность клетки. При определенной величине потенциала на мембране количество ионов К+, пересекающих мембрану в том и другом направлении, оказывается равным, т.е. концентрационный градиент калия уравновешивается имеющимся на мембране потенциалом. Потенциал, при котором диффузионный поток ионов становится равным потоку одноименных ионов, идущих в обратном направлении, называют потенциалом равновесия для данного иона. Для ионов К+ потенциал равновесия равен -90 мВ. В миелинизированных нервных волокнах величина потенциала равновесия для ионов СI- близка к значению мембранного потенциала покоя (-70 мВ). Поэтому, несмотря на то что концентрация ионов СI- снаружи волокна больше, чем внутри его, не отмечается их одностороннего тока в соответствии с градиентом концентраций. В этом случае разность концентраций сбалансирована потенциалом, имеющимся на мембране.
Ион Na+ по градиенту концентраций должен был бы входить внутрь клетки (его потенциал равновесия составляет +60 мВ), и наличие отрицательного заряда внутри клетки не должно было бы препятствовать этому потоку. В этом случае входящий Na+ нейтрализовал бы отрицательные заряды внутри клетки. Однако этого в действительности не происходит, так как мембрана в покое малопроницаема для Na+.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (активный транспорт). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается стремя находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с двумя находящимися вне клетки ионами К+ которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой системе приводит к следующим результатам:
- поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится, активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе, поскольку он сам создает небольшой, но постоянный ток положительных зарядов из клетки, а потому вносит прямой вклад в формирование отрицательного потенциала внутри нее. Однако величина вклада электрогенного насоса в общее значение потенциала покоя обычно невелика и составляет несколько милливольт;
- поддерживается низкая концентрация ионов Na + внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки;
- поддерживая стабильный концентрационный градиент Na + , натрий-калиевый насос способствует сопряженному К+, Na+ -транспорту аминокислот и Сахаров через клеточную мембрану.
Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой проводимостью клеточной мембраны в состоянии покоя для ионов К + , СI-, ионной асимметрией концентраций ионов К + и ионов СI-, работой систем активного транспорта (Na+/K+ -АТФаза), которые создают и поддерживают ионную асимметрию.
Потенциал действия нервного волокна, нервный импульс
Потенциал действия - это кратковременное колебание разности потенциалов мембраны возбудимой клетки, сопровождающееся изменением ее знака заряда.
Потенциал действия является основным специфическим признаком возбуждения. Его регистрация свидетельствует о том, что клетка или ее структуры ответили на воздействие возбуждением. Однако, как уже отмечалось, ПД в некоторых клетках может возникать спонтанно (самопроизвольно). Такие клетки содержатся в водителях ритма сердца, стенках сосудов, нервной системе. ПД используется как носитель информации, передающий ее в виде электрических сигналов (электрическая сигнализации) по афферентным и эфферентным нервным волокнам, проводящей системе сердца, а также для инициирования сокращения мышечных клеток.
Рассмотрим причины и механизм генерации ПД в афферентных нервных волокнах, образующих первично воспринимающие сенсорные рецепторы. Непосредственной причиной возникновения (генерации) ПД в них является рецепторный потенциал.
Если измерять разность потенциалов на мембране ближайшего к нервному окончанию перехвата Ранвье, то в промежутках между воздействиями на капсулу тельца Пачини она остается неизменной (70 мВ), а во время воздействия деполяризуется почти одновременно с деполяризацией рецепторной мембраны нервного окончания.
При увеличении силы давления на тельце Пачини, вызывающей возрастание рецепторного потенциала до 10 мВ, в ближайшем перехвате Ранвье обычно регистрируется быстрое колебание мембранного потенциала, сопровождающееся перезарядкой мембраны — потенциал действия (ПД), или нервный импульс (рис. 2). Если сила давления на тельце возрастет еще больше, амплитуда рецепторного потенциала увеличивается и в нервном окончании генерируется уже ряд потенциалов действия с определенной частотой.

Рис. 2. Схематическое представление механизма преобразования рецепторного потенциала в потенциал действия (нервный импульс) и распространения импульса по нервному волокну
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токов между деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко входу ионов Na+. Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой, равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных импульсов пропорционально амплитуде рецепторного потенциала, а следовательно, силе давления на рецептор. Процесс преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
Природа потенциала действия нервного волокна (нервного импульса)
Мембрана нервного волокна в точке локализации раздражающего электрода отвечает на воздействие очень слабого тока, еще не достигшего порогового значения. Этот ответ получил название локального, а колебание разности потенциалов на мембране — локального потенциала.
Локальный ответ на мембране возбудимой клетки может предшествовать возникновению потенциала действия или возникать как самостоятельный процесс. Он представляет собой кратковременное колебание (деполяризация и реполяризация) потенциала покоя, не сопровождающееся перезарядкой мембраны. Деполяризация мембраны при развитиии локального потенциала обусловлена опережающим входом в аксоплазму ионов Na+, а реполяризация — запаздывающим выходом из аксоплазмы ионов К+.
Если воздействовать на мембрану электрическим током возрастающей силы, то при се величине, называемой пороговой, деполяризация мембраны может достигнуть критического уровня — Е к, при котором происходит открытие быстрых потенциалзависимых натриевых каналов. В результате через них происходит лавинообразно нарастающее поступление в клетку ионов Na+. Вызываемый процесс деполяризации приобретает самоускоряющийся характер, и локальный потенциал перерастает в потенциал действия.
Уже упоминалось, что характерным признаком ПД является кратковременная инверсия (перемена) знака заряда на мембране. Снаружи она на короткое время (0,3-2 мс) становится заряженной отрицательно, а внутри — положительно. Величина инверсии может составлять до 30 мВ, а величина всего потенциала действия — 60-130 мВ (рис. 3).
Таблица. Сравнительная характеристика локального потенциала и потенциала действия
|
Характеристика |
Локальный потенциал |
Потенциал действия |
|
Проводимость |
Распространяется местно, на 1-2 мм с затуханием (декрементом) |
Распространяется без затухания на большие расстояния по всей длине нервного волокна |
|
Закон «силы» |
Подчиняется |
Не подчиняется |
|
Закон «все или ничего» |
Не подчиняется |
Подчиняется |
|
Явление суммации |
Суммируется, возрастает при повторных частых подпороговых раздражениях |
Не суммируется |
|
Величина амплитуды |
||
|
Способность к возбудимости |
Увеличивается |
Уменьшается вплоть до полной невозбудимости (рефрактерность) |
|
Величина раздражителя |
Подпороговая |
Пороговая и сверхпороговая |
Потенциал действия в зависимости от характера изменения зарядов на внутренней поверхности мембраны подразделяют на фазы деполяризации, реполяризации и гиперполяризации мембраны. Деполяризацией называют всю восходящую часть ПД, на которой выделяют участки, соответствующие локальному потенциалу (от уровня Е 0 до Е к ), быстрой деполяризации (от уровня Е к до уровня 0 мВ), инверсии знака заряда (от 0 мВ до пикового значения или начала реполяризации). Реполяризацией называют нисходящую часть ПД, которая отражает процесс восстановления исходной поляризации мембраны. Вначале реполяризация осуществляется быстро, но, приближаясь к уровню Е 0 , скорость се может замедляться и этот участок называют следовой отрицательностью (или следовым отрицательным потенциалом). У некоторых клеток вслед за реполяризацией развивается гиперполяризация (возрастание поляризации мембраны). Ее называют следовым положительным потенциалом.
Начальную высокоамплитудную быстропротекающую часть ПД называют также пик, или спайк. Он включает фазы деполяризации и быстрой реполяризации.
В механизме развития ПД важнейшая роль принадлежит потенциалзависимым ионным каналам и неодновременному увеличению проницаемости клеточной мембраны для ионов Na+ и К+. Так, при действии на клетку электрического тока он вызывает деполяризацию мембраны и, когда заряд мембраны уменьшается до критического уровня (Е к), открываются потенциалзависимые натриевые каналы. Как уже упоминалось,эти каналы образованы встроенными в мембрану белковыми молекулами, внутри которых имеются пора и два воротных механизма. Один из воротных механизмов — активационный обеспечивает (при участии сегмента 4) открытие (активацию) канала при деполяризации мембраны, а второй (при участии внутриклеточной петли между 3-м и 4-м доменами) — его инактивацию, развивающуюся при перезарядке мембраны (рис. 4). Поскольку оба этих механизма быстро изменяют положение ворот канала, то потенциалзависимые натриевые каналы являются быстрыми ионными каналами. Это обстоятельство имеет определяющее значение для генерации ПД в возбудимых тканях и для его проведения по мембранам нервных и мышечных волокон.

Рис. 3. Потенциал действия, его фазы и ионные токи (а, о). Описание в тексте

Рис. 4. Положение ворот и состояние активности потенциалзависимых натриевого и калиевого каналов при различных уровнях поляризации мембраны
Чтобы потенциалзависимый натриевый канал мог пропускать внутрь клетки ионы Na+, необходимо открыть лишь активационные ворота, поскольку инактивационные в условиях покоя открыты. Это и происходит, когда деполяризация мембраны достигает уровня Е к (рис. 3, 4).
Открытие активационных ворот натриевых каналов приводит к лавинообразному вхождению натрия внутрь клетки, движимому действием сил его электрохимического градиента. Поскольку ионы Na+ несут положительный заряд, то они нейтрализуют избыток отрицательных зарядов на внутренней поверхности мембраны, снижают разность потенциалов на мембране и деполяризуют ее. Вскоре ионы Na+ придают внутренней поверхности мембраны избыток положительных зарядов, что сопровождается инверсией (сменой) знака заряда с отрицательного на положительный.
Однако натриевые каналы остаются открытыми лишь около 0,5 мс и через этот промежуток времени от момента начала
ПД закрываются инактивационные ворота, натриевые каналы становятся инактивированными и непроницаемыми для ионов Na+, поступление которых внутрь клетки резко ограничивается.
С момента деполяризации мембраны до уровня Е к наблюдаются также активация калиевых каналов и открытие их ворот для ионов К+. Ионы К+ под действием сил концентрационного градиента выходят из клетки, вынося из нее положительные заряды. Однако воротный механизм калиевых каналов является медленно функционирующим и скорость выхода положительных зарядов с ионами К+ из клетки наружу запаздывает по отношению ко входу ионов Na+. Поток ионов К+, удаляя из клетки избыток положительных зарядов, обусловливает восстановление на мембране исходного распределения зарядов или ее реполяризацию, и на се внутренней стороне через мгновение от момента перезарядки восстанавливается отрицательный заряд.
Возникновение ПД на возбудимых мембранах и последующее восстановление исходного потенциала покоя на мембране оказываются возможными потому, что динамика входа в клетку и выхода из клетки положительных зарядов ионов Na+ и К+ различна. Вход иона Na+ по времени опережает выход иона К+. Если бы эти процессы были равновесными, то разность потенциалов на мембране не изменялась бы. Развитие способности к возбуждению и генерации ПД возбудимыми мышечными и нервными клетками было обусловлено формированием в их мембране двух типов разноскоростных ионных каналов — быстрых натриевых и медленных калиевых.
Для генерации одиночного ПД требуется поступление в клетку относительно небольшого числа ионов Na+, которое не нарушает его распределения вне и внутри клетки. При генерации большого числа ПД распределение ионов по обе стороны мембраны клетки могло бы нарушиться. Однако в нормальных условиях это предотвращается работой Na+, К+ -насоса.
В естественных условиях в нейронах ЦНС потенциал действия первично возникает в области аксонного холмика, в афферентных нейронах — в ближайшем к сенсорному рецептору перехвате Ранвье нервного окончания, т.е. в тех участках мембраны, где имеются быстрые селективные потенциалзависимые натриевые каналы и медленные калиевые каналы. В других типах клеток (например, пейсмекерных, гладких миоцитах) в возникновении ПД играют роль не только натриевые и калиевые, но и кальциевые каналы.
Механизмы восприятия и преобразования в ПД сигналов во вторично чувствующих сенсорных рецепторах отличаются от механизмов, разобранных для первично чувствствующих рецепторов. В этих рецепторах восприятие сигналов осуществляется специализированными нейросенсорными (фоторецепторные, обонятельные) или сенсоэпителиальными (вкусовые, слуховые, вестибулярные) клетками. В каждой из этих чувствительных клеток имеется свой, особый механизм восприятия сигналов. Однако во всех клетках энергия воспринимаемого сигнала (раздражителя) преобразуется в колебание разности потенциалов плазматической мембраны, т.е. в рецепторный потенциал.
Таким образом, ключевым моментом в механизмах преобразования сенсорными клетками воспринимаемых сигналов в рецепторный потенциал является изменение проницаемости ионных каналов в ответ на воздействие. Открытие Na+, Са 2+ , К+ -ионных каналов при восприятии и преобразовании сигнала достигается в этих клетках при участии G-белков, вторых внутриклеточных посредников, связывании с лигандами, фосфорилировании ионных каналов. Как правило, возникший в сенсорных клетках рецепторный потенциал вызывает высвобождение из них в синаптическую щель нейромедиатора, который обеспечивает передачу сигнала на постсинаптическую мембрану афферентного нервного окончания и генерацию на его мембране нервного импульса. Эти процессы подробно описаны в главе, посвященной сенсорным системам.
Потенциал действия может быть охарактеризован амплитудой и продолжительностью, которые для одного и того же нервного волокна остаются одинаковыми при распространении ПД по волокну. Поэтому потенциал действия называют дискретным потенциалом.
Между характером воздействия на сенсорные рецепторы и числом ПД, возникших в афферентном нервном волокне в ответ на воздействие, имеется определенная связь. Она заключается в том, что на большие но силе или продолжительности воздействия в нервном волокне формируется большее число нервных импульсов, т.е. при усилении воздействия в нервную систему будут посылаться от рецептора импульсы большей частоты. Процессы преобразования информации о характере воздействия в частоту и другие параметры нервных импульсов, передаваемых в ЦНС, получили название дискретного кодирования информации.










Фенол - влияние на организм человека
К чему снятся кошки: толкование снов с участием четвероногих пушистиков
Гороскоп рыбы работа деньги на ноябрь
Солянка: особенности национальной кухни
Церковь трех святителей на кулишках Экскурсия храм трех святителей на кулишках